
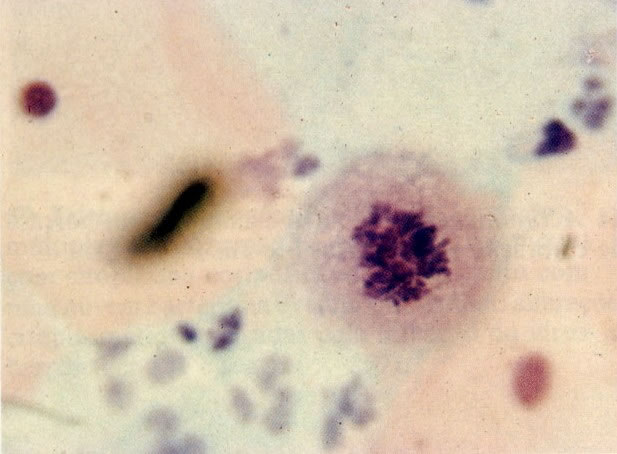
67. Mitoses. Mitoses são menos comumente vistas no material citológico do que em tecido, mas elas podem ser encontradas quando há uma regeneração tecidual rápida. É mais comum achá-la em fragmentos, mas esse tem exemplo é em uma única célula parabasal. Em reações benignas as mitoses são usualmente normais. (X 400)
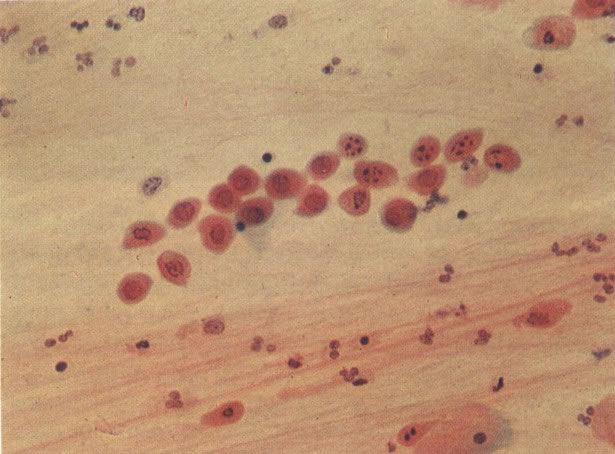
68. Cariorrexe. É necessário distinguir mitoses de fragmentação degenerativa dos núcleos como os vistos nas células parabasais neste campo ponto esta forma de degeneração é descrita como Cariorrexe; outras formas incluem cariólise (69) e citólise (6). (X 120)
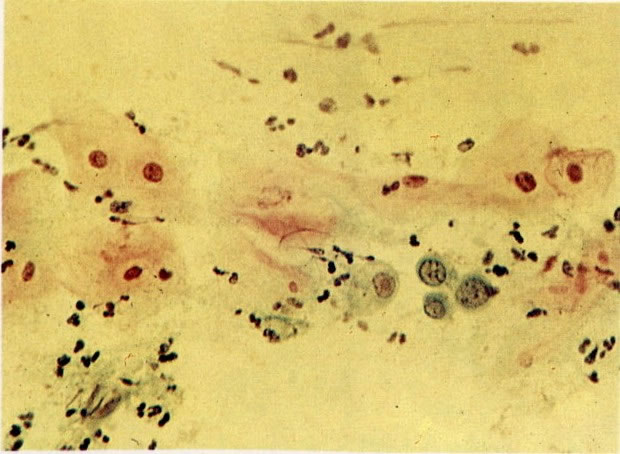
69. Cariólise. O núcleo da célula no centro do campo quase desapareceu. Sua posição é identificada por uma pálida e incompleta membrana.(X 125)
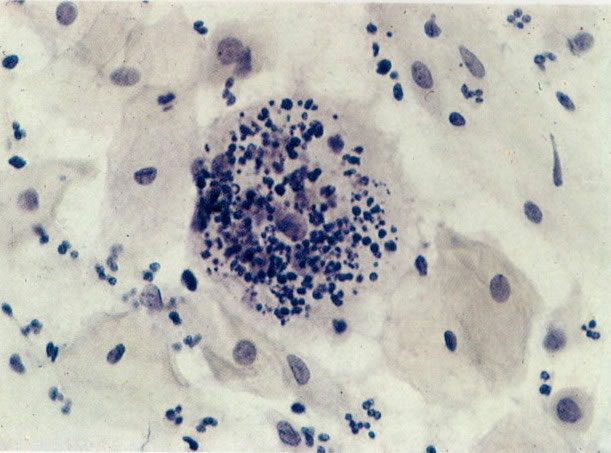
70. Fagocitose. Histiócitos multinucleares gigantes são vistos no esfregaço de pacientes com infecção crônica ou em uma reação granulomatosa. Estes podem fagocitar resto dos nucleares, como visto neste campo. (X160)
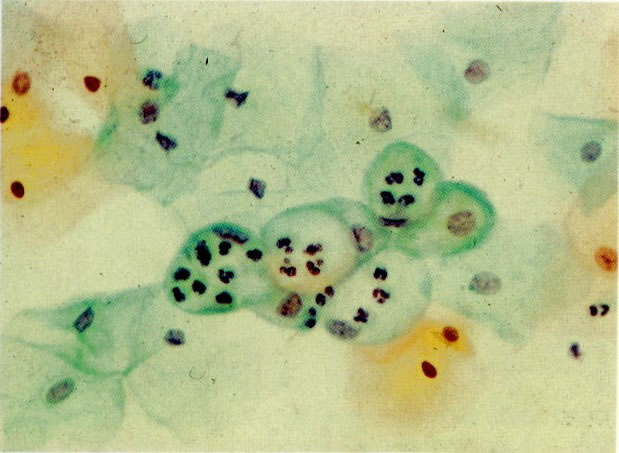
71. Vacúolos citoplasmáticas contendo um polimorfonucleares. Uma aparência lembrando fagocitose é vista quando polimorfos invadem vacúolos degenerativas em células escamosas ou colunares. As células nos vacúolos são algumas vezes referidas como células engolfadas. Notar que neste campo os polimorfos invasores aparecem viáveis quando comparado com fragmentos nucleares em 70.(X 160)
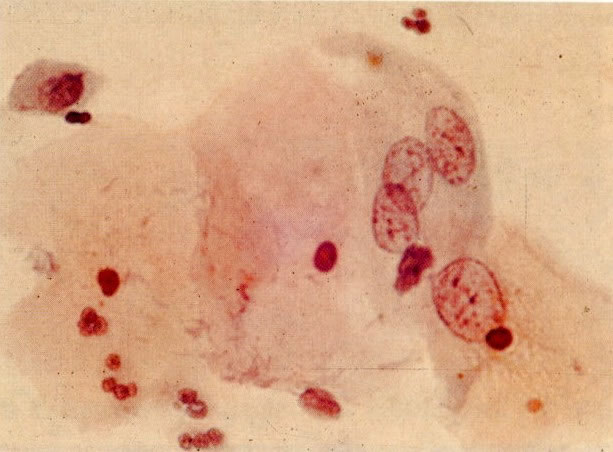
72. Multinucleação. Quando há regeneração tissular rápida, os núcleos podem replicar mais rapidamente que a divisão citoplasmática. Isto resulta em no multinucleação com aspectos nucleares benignos os quais têm aparência idêntica. Neste campo a uma célula trinucleada com núcleos hipocromáticos e aumentados e abaixo desta está uma outra célula com um único no aumentado hipocromático. Mudanças similares têm sido relatadas com deficiência de ácido fólico (van Niekerk. 1966). (X 250)
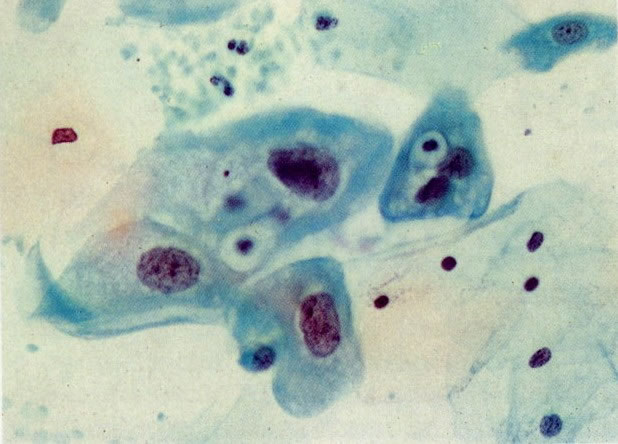
73. Hiperplasia reativa. O aumento dos núcleos é comum nas células que mostram alterações reativas. A cromatina nuclear pode estar dispersa. Sendo assim, o núcleo aparece hipocromático. Em outros casos a degeneração pode causar necrose coagulativa e a cromatina nuclear torna-se hipercromática e pouco nítida. Ambas as aparências nucleares são vistas neste campo. Além disso, halos perinucleares são vistos, juntos com restos, contidos em vacúolos no citoplasma. Uma célula é binucleada.
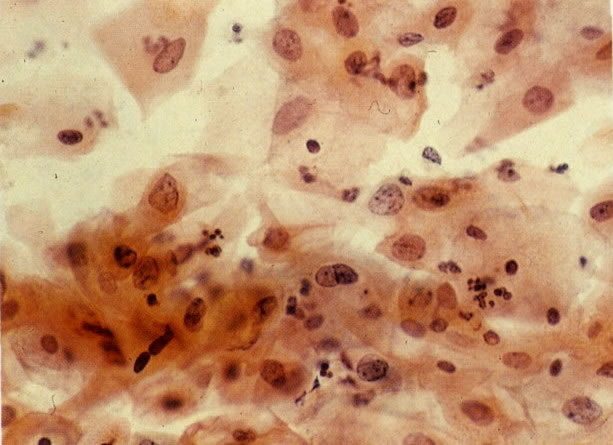
74. Alterações reativas: células escamosas. Neste campo os núcleos são grandes, com dispersão de cromatina dando uma aparência granular. Alguma degeneração nuclear está presente e a coloração citoplasmática é pouco nítida. (X 160)
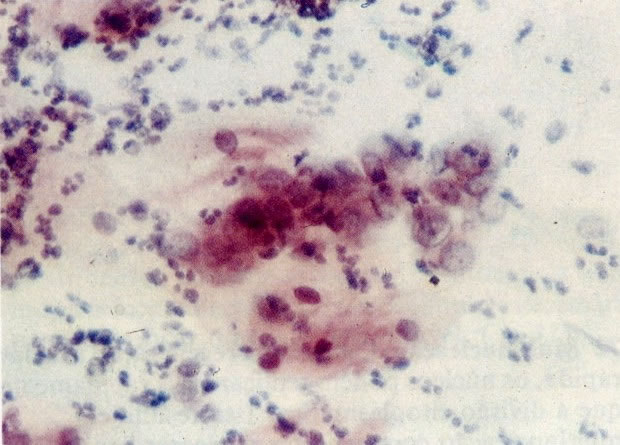
75. Alterações reativas: células colunares endocervicais. Neste campo uma fileira de células colunares endocervicais apresentam-se aumentadas, com núcleos pálidos com nucléolos evidentes. A coloração é geralmente pouco nítida e em algumas células a coloração citoplasmática é anfofílica. Polimorfos estão presentes e muitos estão degenerados. Esfregaços pouco nítidos pobremente corados são vistos comumente com infecção e os relatórios podem ser incertos. Estes esfregaços poderiam ser repetidos após tratamento de infecção. (X 160)
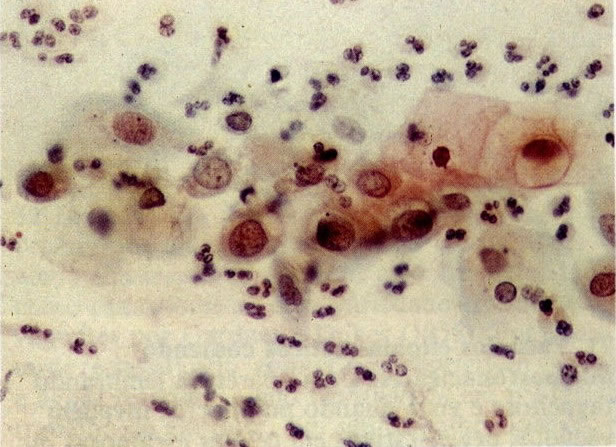
76. Alterações reativas: células metaplásicas. Neste campo de células metaplásicas há núcleos grandes e granulares. Halos perinucleares são vistos em umas poucas células e há algumas vacuolizações citoplasmáticas. Comparações com 156 e 159 mostrarão que quando há aumento da granularidade de cromatina nuclear as alterações começam a ficar próximas da discariose. (X160)
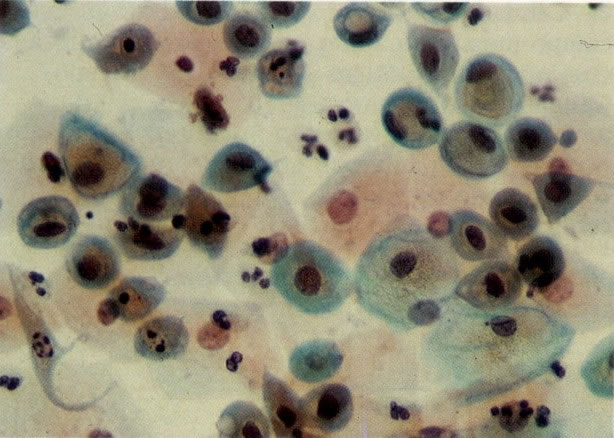
77. Alterações reativas: células metaplásicas. Neste campo há mais degeneração nuclear e hipercromasia; cariorrexe é vista ocasionalmente na célula. O citoplasma é denso e anfólilo em algumas células. (X 160)
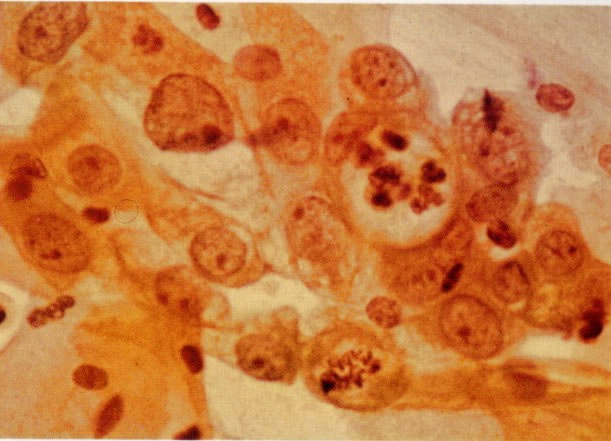
78. Alterações reativas: células colunares endocervicais. Em aumento maior este grupo de células colunares endocervicais mostra alterações reativas acentuadas, com nucléolos proeminentes os quais variam em tamanho e número de célula para célula. Um vacúolo contendo polimorfos está presente em uma mitose normal. Este grau de reatividade pode causar dificuldades ao diagnóstico, mas a presença regular de cromatina finamente granulada seria notada como evidência de que estas células são benignas. (X 250)
[Voltar]
